|
Zero Point Energy and Gigantism in FossilsBarry Setterfield
IntroductionExamining the fossil record is fascinating. It contains a wealth of data, as well as a number of puzzles. One of these is the remarkable size of some of the life forms in the past, not just the famous dinosaurs, but plants, insects, and even a number of mammals as well. The dinosaurs were prominent in the Mesozoic Era. But first let us consider the earlier Paleozoic Era. The flora of the mid to late Paleozoic Era contains large forests of giant club-mosses (Lycopods), giant horse-tail plants (Equisetopsida or Sphenopsids), and huge ferns (Filicopsids). They comprised the majority of the forest fauna that made up the coal measures which formed at the end of the Paleozoic. The Lycopods, whose central genus was Lepidodendron, grew to heights approaching 45 meters. Today, the Lycopods, which we refer to as pendant tassel ferns, attain a maximum size of about 2 meters, but are usually smaller. The Sphenopsids, whose significant species was Calamites, grew up to 18 meters. In comparison, their counterparts, the horsetails, today achieve a much more modest maximum of half a meter. The Filicopsid ferns, such as the tree fern Psaronius, grew 10 meters or more in height. This fern was very similar to the modern Dicksonia or Tasmanian tree fern, whose maximum height today is about 4 meters [1, 2]. We find similar gigantism with the invertebrates that existed during the Paleozoic Era. An important example was reported in New Scientist for the 21 November 2007 [3]. The article stated that, in Germany, strata from the Devonian Period of the Paleozoic Era, yielded the fossilized remains of a giant sea scorpion. The total length of this Eurypterid was conservatively estimated to be 2.5 meters or around 8.3 feet [4]. A number of paleontologists believe that this type of Eurypterid, Jaekelopterus rhenaniae, was the ancestor of modern land scorpions, though some question this conclusion. Since modern scorpions attain a maximum size of about 30 centimeters or about one foot, the size of these early specimens presents a huge puzzle.
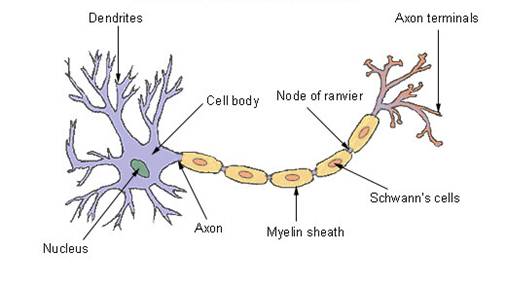
The team, headed by Dr. Simon Braddy, also mentioned that monster millipedes, giant cockroaches, and jumbo dragonflies also appear in this segment of the fossil record. The giant millipede, Arthropleura, was over 2 meters (6.6 feet) long and is considered to be related to the centipedes and millipedes we have today. The largest Paleozoic dragonfly found so far was Meganeura with a wingspans ranging up to 75 centimeters (2.5 feet), and an estimated weight of 450 grams (1 pound). In comparison, the largest dragonfly today has a wingspan of only 7 inches. In fact, Meganeura appears to have been the largest airborne insect of all time [5]. The largest single wing width found measured 33 centimeters, which means a full span would be close to 70 cm. It is not just in the Paleozoic that gigantism can be found among fauna. In Mesozoic strata we have found dinosaurs such as Diplodocus which was 90 feet (27 meters) long, 16 feet (5 meters) tall at the hips, and weighed from 10 to 20 tons. Then there was Apatosaurus which was up to 40 meters (130 feet) long, over 17 meters (55 feet) tall and weighed over 30 tons. As we come into the early Cenozoic, we again see gigantism in a different set of dominant fauna. There was the Megatherium, or giant ground sloth, that weighed 5 tons and, standing on its hind legs and tail, could reach a height of 20 feet. It was apparently one of the largest mammals to walk the earth. In a similar fashion, the giant wombat, Diprotodon, the size of a rhinoceros, may have been the largest marsupial to ever inhabit this planet. In Australia today, wombats have a maximum size under three feet long. In these strata we have also found giant kangaroos, which stood 10 feet (3 meters) tall and weighed 450 pounds (200 kilograms), and an 1100 pound (500 kg) flightless bird, whose fossils have recently been discussed [6]. Gigantism appears to be a significant part of the fossil record, but it is not something we see today. The plants and animals we see in our world are often very much smaller than what appear to be their early relatives which we find only in fossils. At present no satisfactory explanation has been offered for either why things were so large before or, conversely perhaps, why things are so small now. We will first deal with the animals and then with the plants. Problems with Fossil FaunaThe Importance of a Sheath around Nerves Nerves in all vertebrate animals, very much including ourselves, conduct electrical impulses. In the same way a wire must be insulated to keep the electrical current from dissipating, nerves are insulated by a fatty layer called myelin. This myelin coating allows the nerve impulses to travel at a very high speed. This speed rises from about 1 meter per second for a bare nerve fiber, say about 10 microns in diameter, to over 50 meters per second for the same axon sheathed in myelin. A micron is one millionth of a meter. This leads us to a difficulty where some of the giants are concerned. Dr. David Colman, Director of Montreal Neurological Institute and Hospital responded to an article by Sander and Claus in Science, 322 (2007), pp.200-201 where they discussed the means of development of a variety of attributes in dinosaurs. In response, Dr Colman said:
On the figures given in that article, myelinated nerves of the same diameter would require about 1.6 seconds for the round trip reaction time. Dr. Colman continues:
Dr. Colman finalizes by saying,
It is at this point that an important fact need be noted. A.S. Hill et al. state:
Their paper illustrated the difference between the myelinated nerves of the jawed vertebrates and the unmyelinated nerves of the invertebrates (non chordates). This means that invertebrates such as the giant fauna of the Paleozoic, the great sea scorpions, the monster millipedes, the immense dragonflies, had non-myelinated nerves. As will be mentioned below by Dr. Zalc, today there appears to be a limiting size of about 30 centimeters for these organisms [9]. Yet these animals must have been efficient in order to have survived. How was this achieved? The Importance of Bio-Electro-Magnetism
In order to arrive at a solution to this problem, we first need to know some more details about the transmission of nerve impulses. Zalc et al. summarize the situation as follows.
It is thus important to know how the nerve operates. J. Malmivuo and R. Plonsey have made some key comments, which have a bearing on our problem. In [10], page 33, they state:
Malmivuo and Plonsey continue [10],
Then in [10] page 39 they explain,
In other words, if the stimulus is not strong enough, there will be no response. It is only once the stimulus is strong enough to rise above a threshold voltage that any signal will be sent. Thus the voltage difference across the cell membrane initiates an electric current. As early as 1905, Ludvig Hermann suggested that the voltage difference between excited and unexcited regions of the axon would cause small currents to flow between them in such a direction that they stimulate the unexcited region. This has proved to be correct and these currents are now called 'local circuit currents.' More about Axons Malmivuo and Plonsey outline in [10] on page 42 what actually happens as follows:
Malmivuo and Plonsey point out that those two properties are the electrical capacitance per unit length of the axon, and the diameter of the axon. The membrane capacitance per unit length determines the amount of charge required to achieve a certain potential or voltage and therefore the rate of flow of electric current. Large capacitance values mean slower conduction velocities if other factors remain the same. The capacitance is much smaller in myelinated axons than in unmyelinated versions and, as a consequence, the nerve conduction velocity is higher in myelinated axons. The other factor affecting conduction velocity is the diameter of the axon. Signal propagation is linearly dependent upon the diameter of the myelinated axon. In the case of myelinated axons, the nerve impulse propagates by jumping from one node of Ranvier (see diagram) to the next. In other words, the speed of electrical transmission through the nerve depends on both the capacitance and diameter of the axon. How Voltage Differences Arise Between the inside and outside of a cell, there can exist a voltage difference. This occurs because the fluids inside and outside a cell are highly conductive, while the cell's membrane is highly resistive. The voltage change therefore occurs across the width of the cell membrane itself. The voltage difference arises from the action of ionized atoms. The main ions involved with nerve conduction are the sodium Na+ ion and potassium K+ ion. Both of these ions have a positive (+) charge since one electron has been removed from each. The movement of these ions and their concentrations on either side of the cell membrane result in changing voltage. Their movement is achieved through the agency of ion channels, ion pumps and ion transporters, which are embedded in the membrane itself. These agencies maintain different ion concentrations inside and outside the cell. With myelinated nerves, the presence of dense clusters of sodium ion channels on the initial axon segment and at the nodes of Ranvier is key in the production of rapidly conducted electrical impulses. Thus, with nerves, we are speaking about electric charges, voltage differences, and the movement of charges (which constitute an electric current). That means we are dealing with the laws of physics generally and electro-magnetism in particular. The Influence of the Zero Point Energy (ZPE) All electric and magnetic phenomena are dependent upon the strength of a massive field of electro-magnetic energy which pervades the entire universe – the Zero Point Energy [11]. It is called the Zero Point Energy (ZPE) because it has been found to be present even when a vacuum has been cooled to as close to absolute zero as possible. It is made up of all different wavelengths going in every direction at once [12]. All currently accepted cosmological models agree there was an initial expansion of the universe. (Whether or not that expansion continues is not part of this discussion). This initial expansion resulted in an enormous amount of potential energy being invested in the fabric of space in the same way a stretched rubber band possesses potential energy When released, the potential energy in the rubber band will convert to kinetic, or active energy. This conversion process is very rapid at first and then slows down and the potential energy begins to run out. Similarly, the potential energy invested in the fabric of space during its expansion converted into kinetic energy. It appears as this universally pervasive energy we call the Zero Point Energy. And, like the rubber band, the conversion rate from potential to kinetic energy was extremely rapid at first, and then slowed down dramatically [13]. This process of conversion did continue for some time, and while it was continuing, the electro-magnetic ZPE field was becoming stronger as evidenced by astronomical data [14]. The change in the strength of the ZPE, as it built up, had two major effects. First, both the (electric) permittivity and (magnetic) permeability of space increased. Second, it has also been shown that, while electrostatic forces remained unchanged, the field strength of an electric charge is inversely proportional to the square root of the ZPE strength [11]. This same proportionality applies to currents. This also means that when the ZPE strength was lower, all capacitances were also lower in direct proportion. The Zero Point Energy and Biological Effects The result of this, biologically, is that, in the early days of life on earth, when ZPE strength was lower, the field strengths of both Na+ and K+ ions were stronger. Stronger field strengths resulted in stronger currents, as previously mentioned. More importantly, the capacitance of the axons was also lower. It is this which is the determining factor in the rate of signal conduction in the axon. On page 42 of [10], Malmivuo and Plonsey show in equation (2.1) that signal conduction velocity along the axon is inversely proportional to the capacitance of the axon. Mathematically, all other terms which would be variable under changing ZPE conditions cancel out. This leaves capacitance as the sole player where nerve conduction velocities are concerned. Thus, when the ZPE strength was 1/10th of its current value, so also was the capacitance of the axons. Under these conditions, then, the nerve signals were not only stronger, but traveled 10 times as fast down the axon. When data from both astronomical and geological observations are coordinated, it is evident that the strength of the ZPE during the development of life on earth was significantly lower than 1/10 of what it is now. So both nerve signal velocities and reaction times would have been much faster as a result. Under these circumstances, unmyelinated axons would present no problem to invertebrates. The significantly faster nerve signal velocities would allow them to grow to the enormous sizes seen in the fossil record and still be efficient creatures. This also applies to the large dinosaurs. Theoretical problems regarding their reaction times are no longer relevant under these conditions. If, during their time, the ZPE was only 1/10 its current strength, their reaction times would still be only 0.16 seconds. The Role of the Synapse The time-delay in the transmission of nerve signals across the synapse also needs to be considered. The synapse, or synaptic cleft, is the gap between two nerve cells. These synapses also operate as a gateway where signals are passed on to other cells such as muscles or glands. In effect, there is a transmitting terminal on the end of an axon and a receiving terminal at the beginning of the adjacent cell. Nerve signals must travel across this synaptic cleft to get to the next cell or neuron. Today this process may take as long as 1 millisecond. There are two forms of transmission across the synapse: electrical and chemical. Both forms are used throughout the nervous system, but the most common form today is chemical transmission. However, when the synaptic cleft is very small (about 0.0035 microns wide) electrical transmission is more common. This narrow gap allows direct electrical coupling between the neurons. In this case, the signal crosses the cleft essentially instantaneously. In contrast, the synapse depending on chemical transmission is about 0.01 to 0.05 microns, or about 10 times the width of the smaller synapses. Chemical transmission rates are much slower than electrical transmission rates. The question is, has this always been the case? Several steps are involved in the transmission of signals through the nervous system. First, the electrical signal progresses down the axon to its terminal. This terminal is a swelling in the axon and contains neurotransmitter molecules (which include glutamate, acetylcholine, seratonin, histamine, γ-aminiobutyric acid (GABA) and others). The electric signal depolarizes the axon terminal, and this opens a voltage-dependent channel which allows Ca++ calcium ions to flow into the terminal. This causes the terminal to expel the neurotransmitter molecules into the synaptic cleft. These neurotransmitters diffuse across the cleft and then attach to the receptors on the other side of the synaptic gap. This attachment then excites the receiving neuron, which opens a voltage-gated sodium ion channel in response, and so the electric current continues. In the meantime, the neurotransmitter molecules, or "ligands", are expelled from the receptors and recycled back to the axon terminal. Electro-Magnetism in Synapse Diffusion The important point that has emerged from experiments is that the apparently simple diffusion of the neurotransmitter molecule across the synapse, and its subsequent "docking" in the specifically designed receptor, is actually facilitated by electromagnetism. The initial research and experimentation was done by R.F. Stewart and B. M. Craven on the neurotransmitter gamma-aminobutyric acid (GABA). As they examined that molecule in detail, they found that "there is an extensive region of electropositivity around the ammonium group, and an electronegative region around the carboxylate group." It is therefore apparent that this neurotransmitter molecule has a dipole field with a negatively charged end and a positively charged end. This was confirmed by later studies. T. Shikata and K. Hashimoto detailed their experimental results with the neurotransmitters GABA and L-Glutamate [16]. They confirmed that large dipole moments existed with both of these molecules, and that the receptors for these molecules detect differences in both magnitude and direction of these dipole electric fields. They explain, "the receptors effectively control the orientation of GABA and L-Glutamate via dipole-dipole interaction ... i.e. an electric field is generated between the binding sites and the [ligands]."[16] In other words, the apparently simple diffusion of the ligand or neuro-transmitter molecule across the synapse is in fact facilitated by electrostatic attraction, and so, too, is the "docking" procedure. This picture has received a significant boost recently by research performed by P. Kovacic, R. Posos, and C. D. Draskovich. Their key paper regarding this is "Unifying Electrostatic Mechanism for Receptor - Ligand Activity." [17]. They point out that their paper elaborated on an earlier electrostatic mechanism discovered for the receptor-ligand activity. They write
In other words, in all the ligands studied, it has been found that the action of dipoles, electrostatic fields and voltage on both ligands and receptors are responsible for the diffusion across the synapse. Voltage-Driven Diffusion Therefore, even in the case of chemical transmission, we are dealing with electrically driven or voltage-driven diffusion for the transmission of nerve signals across the synapse. We have already noted that the field strength of an electric charge is inversely proportional to the square-root of the strength of the ZPE. This also applies to dipole fields. As a consequence, these ligands would be more strongly attracted to their receptors when the ZPE strength was lower. This attraction, and hence the rate of movement of the diffusing molecule, will be proportional to the voltages involved. Since the voltage from such charges is also inversely proportional to the square-root of ZPE strength, this means the rate of voltage-driven diffusion holds the same proportionality. Therefore, when the ZPE was at 1/10th of its present strength, the rate of diffusion across the synapse was about 3.2 times faster. All biological voltages would change in proportion to this. Conclusion on Faunal Gigantism in Fossils It then appears that the whole process of nerve transmission, including voltage-driven diffusion, may well have been more efficient in earlier geological times when the strength of the ZPE was lower. Other published research of Kovacic, Posos, and Draskovich has indicated that voltage driven diffusion may be operating in a variety of other biological applications. This conclusion is enhanced by the work of Malmivuo and Plonsey [10]. Their studies show that everything from cell behavior to muscular and gland behavior is related to bio-electro-magnetics. A lower ZPE would result, as mentioned, in greater voltage differences, and it is this which would promote faster and more efficient ion transfers, be they within the cells themselves or between cells such as nerves. There are some important ideas to consider in this regard. Both muscular systems and glands, in particular, are dependent upon both the strength of electrical impulses and the associated ion transfers. Bio-electro-magnetics, dealing with both these areas, affects almost every part of an organism’s functioning. Thus, it may well follow that a number of the biological problems we see today would not have been common, or perhaps not even existed, in times past. For instance, if biological systems were more efficient with lower ZPE strengths, resulting in more efficient muscular systems, many heart problems we see today might not have existed before. If glandular activity was also more efficient, immune systems would have also been stronger and more quickly reactive. This type of thing may then indicate a potential for not only gigantism, but longer life spans as well. Cenozoic Extinction of Megafauna The final matter to be dealt with for fossil fauna is the extinction of the megafauna in the Cenozoic. Large mammals were prominent in fossil strata from the Eocene to the end of the Miocene, that is from 55 million down to 5 million atomic years. In the interval from 5 million down to 2.6 million atomic years the extinction of these creatures had became noticeable. During the ice-age the process continued until the last stragglers disappeared in the interval 50,000 to 15,000 atomic years ago. There has been speculation that both human expansion and climate changes were responsible. However, another factor was also at work; the behavior of the ZPE. The curve describing the increase of the ZPE has been shown to be the inverse of the curve that astronomical redshift data follow. This was detailed in a paper presented at the 17th NPA Conference in 2010 by B. J. Setterfield. It is this same astronomical curve that describes the rapidity of transmission of bio-electro-magnetic phenomena since that, too, is inversely related to ZPE strength. One aspect of this common curve now claims our attention. It flattens out with increasing time, but at a fairly high level. Then, as the present era is approached, there is a sudden drop down to its current value. We can pick up this drop in the astronomical data: the redshift curve undergoes a sudden decline as our Local Group of galaxies is approached. The time-frame for this drop starts from about 10 million down to about 3 million atomic years ago. This is in good accord with fossil data. So a key factor in the megafauna extinction was a rapid decline in the efficiency, strength, and propagation rates of bio-electro-magnetic effects. This decline occurred as the ZPE increased to its current level. Problems with Fossil FloraBecause flora were so prolific in both size and numbers in the fossil record, it may well be that some factor or factors promoting plant growth have changed. To understand possible causes for any change in plant size and numbers, we must examine the basic processes involved in plant growth. This means we must examine both photosynthesis in plants, and the behavior of light with time given a lower ZPE strength in the past. Let us start by looking at light itself. Light through Time A photon of light is released when an electron on any atom, from any element, is forced out of position relative to the nucleus and then snaps back into position. The energy it took to force it out of position is the energy released as a photon of light when it snaps back into position. Data indicate that the speed of light was much faster in the past [18, 19]. It was subsequently demonstrated that the speed of light is inversely dependent upon the strength of the massive field of electro-magnetic energy which pervades the entire universe, namely the Zero Point Energy (ZPE) [12]. The increase in the strength of the ZPE through time has had two effects on light. First of all, a stronger ZPE slowed light down. Since ZPE waves go in all directions, they impact each other in somewhat the same way as the wake of a speedboat impacts waves in the ocean. Where waves meet, they crest and form whitecaps which die down quickly. When ZPE waves meet, they create a concentration of energy that results in the formation of a positive and negative pair of particles, which flash into existence momentarily, then re-combine and annihilate. For this reason they are referred to as virtual particles. These particles can absorb a photon of light the same way any other particle can. However their almost instantaneous recombination releases the photon to continue on its way. The more virtual particles a photon of light must navigate, the longer it takes to reach its final destination, or point of final absorption. It has been estimated that today, at any given instant, there are about 1042 virtual particles flashing into and out of existence in any given cubic meter. Consequently, light photons encounter huge numbers of these virtual particles even over very short distances. This means that when the ZPE was weaker, there were fewer virtual particles at any one time in any given volume to slow down the passage of a photon of light. As the ZPE built up, then, the speed of light necessarily slowed. Effectively, the vacuum has become "thicker" with these virtual particles over time. This means that before, with a lower ZPE, and hence fewer virtual particles, the speed of light was higher and therefore more photons of light were arriving at any location at any given instant. It has also be shown that light generation processes in stars, such as our sun, were correspondingly more rapid [20]. Higher Speed Also Means Softer Light With greater numbers of waves or photons reaching them, it might be thought that plants and trees would burn up. However, this is not the case. To understand why, it is necessary to understand something about waves in motion. A basic formula applies in such cases. It states that, for any given wave, the square of the wave amplitude (or height) multiplied by the wave velocity is a constant [21]. So initially, when the speed (the velocity) of light was high, the heights or amplitudes of these light waves had to be quite small. As the speed of light reduced, the heights or amplitudes of these light waves increased. A similar situation can be seen in a tsunami wave. When traveling fast in the middle of the ocean, it may only be a meter high. As it nears land, the changes it encounters simultaneously reduce its speed and increase its height. The brightness, or intensity, of light is determined by the wave amplitude (height). The amplitude changes with the speed. When the speed of light was higher, the amplitude was lower – thus the light per photon from the sun was less bright. But there were proportionally more photons arriving per second with this lower amplitude. The result was that the total brightness would have been the same as now. When light speed slowed, amplitude increased. Today, the light from each photon is very bright, but there are fewer photons arriving per second. The result, though, is that the overall brightness has not changed. It is important to note that the changes in the ZPE, and therefore in the amplitude and velocity of the light, did not alter the wave lengths themselves; they were the same from maximum point to maximum point [12]. This means the colors were not different, as colors are a result of wave length not wave height or speed. The height of the light wave indicates its brightness and it seemed intuitive for a long time to associate the brightness with the energy of the light itself. In the early twentieth century, however, something called the photoelectric effect was discovered. It was already known that if a ray of light hit a bit of metal, an electron could be discharged and start an electric current. However, there was a surprise involved: some light, no matter how brightly it was concentrated on the metal, failed to activate an electric current. On the other hand, certain colors of light, no matter how faint, did activate the current. Light color is determined by wavelength, and therefore is the determining factor in the amount of light energy any light wave contains. The energy is not a matter of brightness, but of color. As far as the early plants were concerned, the outcome was that the strength of each photon’s impact on a leaf was less, even though its energy (color) was the same as now. So while there were more photons hitting each leaf at any given time, it was like something being bathed in this soft light. Today the overall brightness of the light is the same. However, instead of the bathing effect, today’s light is more like hailstones hitting: fewer, but harder, even though the same amount of water is involved. In summary, when data is coupled with physics, the indication is that plants received more photons of light of a given energy per unit area per second than they do now. But the overall brightness of the total number of photons received per second has remained unchanged. Given this, we need to consider what photosynthesis really does. Overview of Photosynthesis Photosynthesis is the basic process whereby plants, algae, and some bacteria use energy from sunlight to convert water and carbon-dioxide to oxygen and sugars. A basic equation which describes the overall process is as follows:
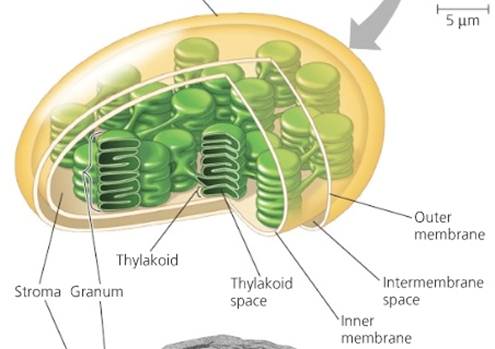
The "Light Reactions" One basic difference between animal cells and plant cells is the existence of something called chloroplasts in the plant cells. In plants and algae, (the eukaryotes), the light reactions all occur within the chloroplast. The chloroplast is a specialized component of certain cells which not only contains chlorophyll but also the systems responsible for photosynthesis (see Fig.2). The matrix of the chloroplast is a thick fluid where various ions, enzymes and molecules are found. This matrix is called the stroma. Bacteria (prokaryotes) have a slightly different arrangement but with a similar purpose.
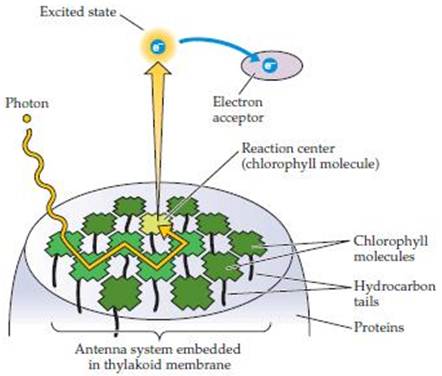
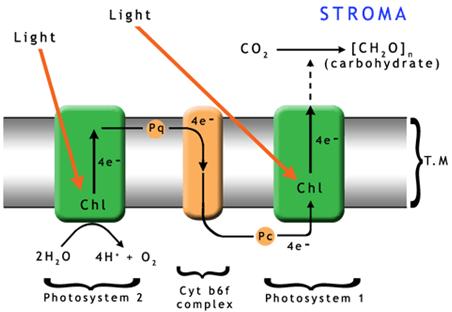
Processes inside a Chloroplast The process begins when light photons are collected and absorbed by the surface ‘antenna pigments’ (see Fig.3). These pigments contain chlorophyll, but they do not use the light energy themselves. They collect the light energy and transfer it deeper into the plant cell where a series of pigment molecules and proteins together, called the ‘reaction center,’ begin the actual energy conversion in the thylakoid membranes. The antenna pigments, are made up of chlorophyll a and chlorophyll b as well as some carotenoid molecules. Chlorophylls aand b differ in the wavelengths they absorb best, but both absorb red and blue much better than yellow and green. However, the carotenoids fill this gap by preferentially absorbing green light. The entire antenna complex, comprising from 100 to 5000 molecules, ensures that the majority of light photons will be trapped and passed on to their associated reaction centers. The number of pigment molecules in the antenna complex is flexible. It is usual for systems growing in low light or shady conditions to have more antenna molecules connected to each reaction center than those in full sun. The process by which energy is passed down to the thylakoid reaction centers is via electron transfer. There are two photosystems with which the antenna complexes and their reaction centers are associated. In Photosystem II (PS II) the reaction center contains chlorophyll P680 while Photosystem I (PS I) contains chlorophyll P700. Absorption of one photon of light by PS II removes one electron from P680. With its resulting positive charge, P680 can remove one electron from a molecule of water. When this process has occurred four times (using two molecules of water), one molecule of oxygen is released and the hydrogen has been split into four electrons and four protons. This may be described by the following equation:
The conversion of ADP (the Di-Phosphate form) to ATP (the Tri-Phosphate) involves the addition of one more phosphate group to the original molecule. The energy required to attach this third phosphate is where the energy is stored. Finally, electrons are transferred via Photosystem I to NADP+ to make NADPH, which is the reduced form of nicotinamide adenine dinucleotide phosphate. Again, the addition of the hydrogen requires energy, and it is in this bonding that the energy is stored for the cell to use later. The two products resulting from these photon driven reactions are therefore NADPH and ATP, both of which supply energy for metabolic processes, and, as a result, are sometimes called the energy currency for cells. The rate of energy transfer between antenna pigments on the way to the reaction centers is very rapid, ranging from 10-15 seconds down to 10-9 seconds. In the reaction centers, the charge separation reaction occurs in times that typically range from 3 x 10-12 to 3 x 10-11 seconds. The synthesis of ATP and NADPH is much slower, taking from 10-6 down to 2 x 10-3 seconds [22]. This means that the electron transport chain is fast enough to utilize the majority of the incoming photons. However, in today’s full sunlight the formation of ATP and NADPH cannot keep up with the amount of light the plant absorbs. This is not due to the amount of light received, but to the speed of the reactions. The result is photo-inhibition and results in the plant shutting down part of the electron transport chain. Photo-Inhibition Normally, the chlorophyll molecule itself has two electrons in its outer shell, and they will be spinning in opposite directions. More light than the plant can handle tends to cause the formation of "triplet state" chlorophyll. In this state, the two electrons in the outer shell have identical spin orientations instead of opposite spins. This triplet chlorophyll readily reacts with oxygen which leads to a very reactive single oxygen atom being released (these are called free radicals in our own bodies). These oxygen atoms damage proteins. This damage then slows down the formation of ATP and NADPH. However, experiments have shown the problem can be countered when carotenoids are in close proximity to the chlorophylls. This counter-action occurs because carotenoids efficiently "quench" the triplet states of chlorophyll and so avoid the formation of the singlet, or free radical, oxygen. Carotenoids are fat-soluble pigments found primarily in plants. They are critical to the photosynthesis processes. Carotenoids are anti-oxidants, protecting against not only free radical oxygen atoms, but over-exposure to light as well. Plants produce carotenoids in response to exposure to light; the more the exposure, the more the carotenoid production occurs up to what appear to be certain limits [23, 24]. There is no reason to doubt that the same protection which is afforded plants today would also have been in operation in times past. In support of this, it has been shown through experiments and research that ATP and NADPH production will increase when more light photons are available. When we put this information together, we are left with the interesting hypothesis that, in times past, when the speed of light was faster, the increased number of light photons reaching plants was prevented from doing possible damage due to increased carotenoid production. At the same time, the plants’ abilities to store more energy were increased. As the speed of light decreased with time, plants received fewer photons of light per unit of time and carotenoid production dropped proportionately, as did the ability to store energy via the production of ATP and NADPH. The Carbon Fixation Process Carbon fixation through the Calvin cycle is the second phase of photosynthesis and is sometimes labeled the “dark phase,” or "dark reactions". These reactions take carbon dioxide from the air and, using the energy stored in ATP and NADPH, convert the energy to sugars. This cycle uses a catalyst which changes carbon dioxide into two identical molecules each containing 3-carbon atoms. These 3-carbon molecules are then the basis for the synthesis of glucose and other sugars. Because there are 3-carbon atoms in the molecules produced by the catalyst, this process is referred to as the C3 pathway. The catalyst that is used in this process is the enzyme ribulose biphosphate carboxylase oxygenase, which is usually referred to (for obvious reasons) as Rubisco. Rubisco is capable of catalyzing two different reactions. The Calvin cycle, as explained above, is the primary reaction using Rubisco. In it carbon dioxide is converted to sugars. However, when carbon dioxide concentrations are low, Rubisco enables a different reaction by taking available oxygen and, combining it with the plant’s carbon, producing carbon dioxide. This is called photorespiration, and effectively reverses all the hard work the plant has done using carbon dioxide to form sugars. In order to avoid robbing the plant of its own sugars, relatively high carbon dioxide concentrations are desirable so that reactions proceed along the usual C3 pathway instead of reverting to photorespiration. Interestingly, the fossil record indicates that carbon dioxide concentrations were about 0.3% of the total atmospheric content at the time of plant gigantism. This is about ten times the amount in our atmosphere today (which is 0.025%). When carbon dioxide was at that high level, photorespiration would have been extremely rare. This, then, is one part of the explanation for the gigantism in fossil plants. The carbon dioxide content remained at 10 times its current level throughout the Paleozoic and Mesozoic Eras. But shortly after the onset of the Cenozoic Era, nearly 60 million atomic years ago, the CO2 concentration in the atmosphere began dropping until 20 million atomic years ago when it reached today's concentration. About the time that the CO2 concentration dropped, about 10,000 species of flowering plants, or angiosperms, in 19 different families became prominent, including some 4500 species of grasses, or Poaceae. These plants all have a mechanism which increases their CO2 concentrations before the Rubisco catalysis occurs, enabling them to thrive in a low CO2 content. This may be a major reason the angiosperms became so prominent in the Cenozoic. These angiosperms employ a supplementary method of CO2 uptake in addition to the usual C3 pathway. This method produces oxaloacetic, malic and aspartic acids which all contain 4-carbon molecules. As a consequence, these plants are called C4 plants. They have structural changes in their leaf anatomy so that their C3 and C4 pathways are separated to allow their reactions to occur in different parts of the leaf. In this context, the situation with the horsetails comes to the fore. These show up as giants in fossils. But they have so little leaf surface to capture the light and provide energy to the plant; how did they grow so tall? The answer may be hinted at in an article by Hibberd and Quick in Nature [25]. In that article, they report some C3 plants, notably celery and tobacco, also had a C4 type of mechanism for fixing carbon in their stems and leaf stalks. In this case, the CO2 from root respiration diffuses into the sap at concentrations considerably higher than air. The CO2 rises up in the plant, and is fixed, using Rubisco, by this C4 mechanism, in the stems and leaf stalks rather than diffusing out to the atmosphere. This occurs in the exact positions that would allow carbon fixation and lignin synthesis in the early horsetails and club mosses. Hibberd and Quick’s data also show that this process occurs at a high rate because the production of the intermediate PEP molecule in the veins was 18 times greater than in the leaves. The indication was that this may be a more general phenomenon than otherwise expected. The reason is that large quantities of light are needed for this mechanism to activate and begin operation. Since there would be an abundance of light photons in the Paleozoic Era due to a lower ZPE, then we have a very plausible explanation for the size of club mosses and horsetails in the fossil record. Activating the Rubisco Enzyme As noted, "light reactions" all proceed with rapidity without photo-inhibition provided that the concentration of carotenoids is maintained at a suitable level. When the flux of photons from the sun was higher, the production rate of carotenoids, ATP and NADPH would have been correspondingly greater, in line with what our experiments today show. But the carbon fixation reactions of the Calvin cycle, which use ATP and NADPH, only export stable products to the plant in times ranging from 10-3 seconds down about 0.3 seconds. Examination of these reactions indicates that the rate limiting step is the performance of the Rubisco enzyme, which is readily de-activated. Since the catalysis itself is rapid, it appears that the problem is to get the Rubisco enzyme activated so that catalysis can occur. Up until 1987 it had been thought that the absence of either or both the Mg++ ion and a special carbamate group was the cause of Rubisco deactivation. But then it was found that the enzyme rubisco activase was vital to the process of activation. It was then verified that all the other problems with activation of the Rubisco enzyme were secondary, and that activation depended entirely on the concentrations of both Rubisco and rubisco activase. Several important conclusions also came from two other studies [26, 27]. In the first, it is stated that the presence of ATP leads to Rubisco activation through the action of rubisco activase. Since ATP is a product of the "light reactions", its concentration would have been higher long ago, when photon reception rates were higher. Thus Rubisco activation would also have been higher. The second conclusion from the follow-up study involves a data-based equation for the rate of Rubisco activity which stated that
The quantities inside the square brackets are the concentrations with k a proportionality constant. Therefore, based on experimental data, this means that when the rubisco activase concentrations are higher, so are the activation rates. It is in this context that the authors of the follow-up report noted that there was only one rubisco activase site for every 72 Rubisco sites. However, as mentioned, rubisco activase is directly dependent upon the amount of ATP produced. The increased number of photons available to the plant at any one time in the past would have not only increased production of ATP and therefore the activity of the rubisco activase, it may well have meant there were more rubisco activase sites per Rubisco sites. This combination would have made for a much more efficient series of processes, which would then contribute to a greater reaction rate and thus more plant growth. Changes in the Rubisco Enzyme There is another clue which needs to be followed. Today we have identified three forms of Rubisco plus a Rubisco-like protein (RLP). Are they descendents of an original form of Rubisco, or have all four always been present? Each of today's forms is missing something that one of the other forms has. In the same way we see speciation in animals, perhaps Rubisco itself has degenerated into four sub-forms which started as a much more complete and efficient original. Indeed, this is not an unusual suggestion. F. R. Tabita et al. noted in their abstract that
This indicates that there was probably an ancestor from which the three Rubiscos and Rubisco-like proteins were derived which was free from the problems that Rubisco currently has. Other Implications for the Fossil Record These conclusions have implications in other areas as well. For example, in the ocean, phytoplankton also fix carbon by photosynthesis. These plankton are made up of autotrophic, prokaryotic, and eukaryotic algae such as diatoms, cyanobacteria, dinoflagelletes, and coccolithophores which inhabit the near surface regions of the ocean. Because photosynthesis plays an important role in the growth and reproduction of these algae, it might be expected that all these forms would proliferate under conditions of both higher photon counts and more available carbon dioxide. This in turn would support larger numbers of zooplankton which feed on the phytoplankton. Scientists working for the Census of Marine Life reported in a News Release for 4th May, 2006, that zooplankton represent a fundamental link in the food chain between ocean plant life and larger organisms. Literally thousands of tiny animal species are involved. Consequently, since the food supply may well have been more plentiful in the past, larger populations of marine species could have been supported. It might also be noted that the bacterio-plankton are involved in re-mineralizing the water, so this process would have been enhanced as well. A study by M. Mitra and A Melis mentions an additional factor. They reported that experiments with algae have shown that, under high photon counts, chlorophyll antenna sizes in algae will individually reduce. When this happened, it was found that there was greater transmittance of light deeper into the culture, photo inhibition was reduced, and efficiency significantly improved [29]. Under conditions of higher photon counts, this process would assist algae to utilize all available photons without photo-inhibition. To give some idea of the consequences of this in earlier times, consider the Paleozoic trilobites. Many are described as either filter feeders, or pelagic planktonivores, or considered to have grazed on beds of algae. These are the very food items that would have proliferated due to a high photon count. As a consequence, the huge colonies of trilobites which appear in the fossil record no longer present an enigma, but rather are a testimony, along with Paleozoic forests, of an much higher photon reception rate as well as a higher carbon dioxide content. Indeed, the whole ecosystem would have been more vigorous and robust. Since this ecosystem is the basis of the oceanic food chain, it might be expected that the numbers of ocean creatures would also have been higher. The Growth of Stromatolites Stromatolite fossils may give us another good example. It is true that some of what have been identified as stromatolites are not, but are rather abiotic. However analysis has shown that many are formed by cyanobacteria (blue-green algae). Stromatolites are still growing in Shark Bay, Western Australia and the Exuma Cays in the Bahamas. These structures may be domed or branching or vase-shaped, or a variety of similar mound-like shapes. The algae use water, carbon dioxide and sunlight to produce oxygen and calcium carbonate. A layer of mucus forms over mats of cyanobacterial cells. In these mats, sand, grit and other debris from the surrounding area are trapped in the mucus and become cemented with calcium carbonate. As a result, the structure is built up of thin laminations of limestone. Stromatolites first appear in the Precambrian, with the earliest confirmed microbial structure dating at 2.73 billion atomic years ago [30]. Their growth rates today have been used as markers of the time needed for their sizes in the fossil record. Once again, however, photosynthesis plays an important part in the growth and reproduction of the algae making up most stromatolites. Consequently, higher photon counts and more carbon dioxide would enhance their growth rate as well as their numbers. For this reason, the use of stromatolite colonies in the fossil record as markers of a long growing time, and hence an equally long interval without deposition, may have to be reassessed. Chalk Formation Microscopic green algae, known as coccolithophores, are also photosynthesizers. They also would grow more rapidly and proliferate with the higher photon count when the ZPE was low. These coccoliths are the main constituents of the Upper Cretaceous chalk beds in the UK and across western Europe. In England, the Chalk beds can total a thickness of 215 meters (700 feet) and they form the White Cliffs of Dover. The coccoliths make calcium carbonate shells which are the basic constituent of these chalk beds. However, the environment which allowed this massive amount of chalk to be formed has always been a puzzle. This study offers a potential solution. ConclusionThe gigantism we see in fossils has always puzzled people. Was there something genetic going on, or was it something to do with the environment itself? As we have examined some material in this paper, we have looked at environmental causes, with only a moment’s question regarding the genetics of Rubisco itself. But it looks as though we can attribute most of what we see in the gigantism in fossils to environmental effects. Because ZPE strength was lower in the past, the electric and magnetic properties of the vacuum were different from now. This also meant that voltages and currents were inherently stronger and that electric and magnetic interactions were much faster. Because chemical reactions in both plants and animals are based upon some form of electric (ion) transfers, they react to the properties of space itself, and thus to the Zero Point Energy. So when the ZPE itself was lower, the chemical/electrical processes were both faster and more efficient. This means that the lack of myelin on the nerves of reptiles, for instance, did not stand in the way of their continued growth, as nerve impulses were much faster and capacitances proportionately lower, allowing the animal to respond quickly to stimuli. A lower ZPE also meant there were more light photons arriving per second at a lower amplitude. So although the overall brightness was constant, and the photon energy (color) was unchanged, photosynthesis itself was more efficient. The outcome was that flora could grow larger and more prolifically. Thus, while genetic factors may also have been involved, there is strong evidence that it was the environment itself which was primarily responsible for the gigantism we see in the fossil record. References[1] M. E. White, The Greening of Gondwana, p. 75, (Australia: Reed Books, 1988). * All websites and DOIs were retrieved 3 March 2011.
|